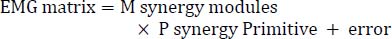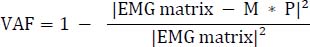|
|
|
| |
| ABSTRACT |
|
This study compared the neuromuscular control strategies and force-time profiles of three upper-limb pushing tasks-the standard push-up (SP), plyometric push-up (PP), and standard squat push-up (SSP)-representing the endurance-, power-, and strength-oriented continuum of upper-limb pushing performance. Fifteen male rugby athletes performed SP, PP, and SSP on dual force plates while surface electromyography (EMG) (12 muscles) and vertical forces were concurrently recorded. EMG signals were processed and decomposed using nonnegative matrix factorization to extract muscle synergies. Synergy modules were evaluated using cosine similarity and paired-samples t tests. One-dimensional Statistical Parametric Mapping (SPM1D) was employed to compare synergy primitives and force-time profiles across tasks. Two synergies reconstructed all tasks (VAF > 0.95). Synergy modules differed among conditions (cosine similarity < 0.90), with task-specific changes mainly involving distal forearm, scapular-trunk, and elbow-extensor muscle weightings. SPM1D identified task-specific differences in synergy primitive 1 during 0%-12% and 48% - 71%, and in primitive 2 during 71% - 100%, with PP and SSP exhibiting higher late-phase activation. PP generated higher force than SP during 13% - 79%, while SSP produced higher force during 33% - 36% and 56% - 83%, but lower force early and near takeoff. Despite sharing two synergies, the three pushing tasks exhibited distinct synergy structures, activation timing, and force-time profiles. PP emphasized rapid early-to-mid phase propulsion, whereas SSP relied on sustained late-phase force. These findings demonstrate task-specific neuromechanical regulation and may help inform exercise selection for upper-limb strength and power development in trained athletic populations. |
| Key words:
Electromyography, explosive movements, neuromuscular coordination, motor control, time-series analysis
|
Key
Points
- Although SP, PP, and SSP were all reconstructed by two muscle synergies, their synergy structures and activation timing differed clearly, indicating task-specific reorganization of upper-limb neuromuscular coordination.
- PP and SSP produced greater concentric force than SP over substantial portions of the movement cycle, but they showed distinct force-generation strategies: PP emphasized rapid early-to-mid phase propulsion, whereas SSP relied more on sustained late-phase force output.
- These findings demonstrate that similar pushing movements can be achieved through different neuromechanical regulation strategies, which may inform exercise selection for upper-limb strength and explosive power development in trained athletic populations.
|
Upper-limb strength and explosive power are pivotal determinants of performance across a wide range of competitive sports (Nichols and Szivak, 2021), particularly in tasks requiring rapid high-force output, such as upper-body collisions and passing in basketball, offensive drives and blocking in rugby, and various throwing actions (Dhahbi et al., 2017b; Koch et al., 2012; Speranza et al., 2018). In these sport-specific contexts, athletes must rapidly coordinate multiple joints and muscle groups to produce high-magnitude force impulses within a strictly limited time window (Baiget et al., 2021; Lum et al., 2022). Owing to its versatility, ease of standardization, and ability to impose diverse mechanical demands, the push-up is widely used both for upper-body strength training and for assessing upper-limb strength, power, and neuromuscular function (Dhahbi et al., 2022; Rogers et al., 2020). Different push-up variants emphasize distinct components of upper-body pushing ability. The standard push-up (SP) is commonly used to assess muscular endurance (Ning and Sheykhlouvand, 2025). The plyometric push-up (PP) evaluates explosive strength (Zalleg et al., 2020), and its peak force has been associated with tackling performance in ball sports (Speranza et al., 2018). In contrast, the standard squat push-up (SSP) produces mean forces that strongly predict 1RM bench-press performance and therefore serves as a practical method for assessing upper-body strength and power (Wang et al., 2017). Previous investigations have primarily relied on discrete outcome variables-such as peak force, rate of force development (RFD), mean power, and electromyography (EMG) amplitude-to quantify performance during different push-up tasks (Dhahbi et al., 2017b; García-Massó et al., 2011; Wang et al., 2017). Consistently, PP has been shown to elicit markedly greater peak ground reaction forces (GRF) and higher RFD compared with SSP (Dhahbi et al., 2017b), reflecting its heightened explosive demand. Such differences may be partly explained by the stretch-shortening cycle (SSC), a muscle-tendon mechanism that enhances force production and improves energy efficiency compared with isolated concentric actions (Nicol et al., 2006). However, these conventional metrics capture only instantaneous or phase-specific outputs and are insufficient to characterize the temporal dynamics of force generation or its underlying neuromuscular regulation. In reality, force production is not instantaneous but emerges from continuously modulated neuromuscular activation over time (Del Vecchio, 2023), highlighting the need for analytical approaches capable of capturing these temporal coordination processes. In recent years, muscle synergy analysis has emerged as a robust analytical framework for examining how the central nervous system (CNS) organizes multi-muscle coordination during complex movements (Fan et al., 2024; Frère and Hug, 2012). By decomposing multi-channel EMG into low-dimensional synergy modules and temporal activation primitives, this method elucidates how muscles are coordinated through task-specific timing and weighted contributions. Prior evidence shows that kneeling PP elicit higher activation of the pectoralis major, triceps brachii, and anterior deltoid compared with traditional PP (García-Massó et al., 2011), highlighting the influence of task intensity on upper-limb neuromuscular demand. Moreover, although the number of synergy structures is largely conserved across push-up tasks performed with different hand positions, the associated synergy weights and activation timings vary systematically with task intensity and postural-control demands (Fan et al., 2025). Collectively, these findings indicate that task-specific control of pushing actions may be reflected not only in changes in EMG amplitude but also in adjustments of synergy structures and temporal activation patterns. Therefore, examining synergy-level organization across different push-up variants may provide deeper insights into the neuromechanical mechanisms underlying force generation during pushing movements. This can provide a foundation for developing training and rehabilitation strategies grounded in physiological principles. Accordingly, the present study examined three representative pushing modalities (SP, PP, and SSP) to investigate task-specific differences in upper-limb synergy organization and dynamic characteristics of force generation during the concentric phase. We hypothesized that: (1) the three pushing tasks would exhibit distinct time-dependent force profiles during concentric propulsion; and (2) each task would demonstrate clear task-specific differences in synergy module composition and activation timing. By elucidating the neuromechanical coupling that governs concentric force production, this study aims to provide mechanistic evidence that may inform upper-limb force training and movement-control assessment in trained athletic populations exposed to high-intensity upper-limb loading. SubjectsFifteen healthy, trained male rugby athletes (age: 21 ± 1.8 years; height: 1.83 ± 0.05 m; body mass: 82 ± 5.6 kg) voluntarily participated in this study (McKay et al., 2021). A sample-size justification was conducted using G*Power 3.1 for a one-way repeated-measures ANOVA with three measurements. Assuming a medium effect size f = 0.25, α = 0.05, power = 0.80, one group, and a correlation among repeated measures of 0.75 based on the average inter-condition correlation observed in the present dataset, the required total sample size was 15. All participants were right-handed and had completed at least five years of structured sport-specific training, ensuring adequate proficiency in upper-limb pushing tasks. None had a history of upper-limb surgery, and no participant reported musculoskeletal injury, neurological disorders, or pain affecting upper-limb pushing movements within the preceding six months. Prior to data collection, all participants provided written informed consent after receiving a comprehensive explanation of the study procedures, potential risks, and their right to withdraw voluntarily at any time. All procedures were conducted in accordance with the Declaration of Helsinki (1975, revised 2013) and were approved by the Jeonbuk University Ethics Committee (JBNU2024-09-015-002).
DesignThis study employed a cross-sectional, within-subject repeated-measures design to compare neuromuscular control strategies and force-time characteristics across three upper-limb pushing tasks (SP, PP, and SSP). All participants performed the three tasks in a randomized order. Each task was executed on dual force plates to quantify bilateral GRF, while kinematic and EMG signals were simultaneously recorded to assess movement events and neuromuscular coordination.
MethodologyAll testing was conducted in a controlled biomechanics laboratory between 15:00 and 17:00 to minimize potential circadian effects on neuromuscular performance. Participants were instructed to avoid strenuous upper-body exercise and alcohol for 48 h, caffeine for 8 h, and food intake for 2 h before testing. Before testing, participants completed a standardized warm-up consisting of 5 min of light jogging, 5 min of static and dynamic upper-limb stretching, and task familiarization for all three pushing tasks. A 5-min seated rest was provided to ensure a return to physiological baseline before data collection. Each participant performed the three upper-limb pushing tasks (SP, PP, and SSP) in a randomized order to minimize order effects. For all tasks, participants placed their hands on two independent force plates, maintained a neutral trunk alignment, and avoided compensatory movements. Hand width on the force plates was standardized in the prone starting position by aligning the medial borders of the hands with the vertical projection of each acromion using a plumb line (Fan et al., 2025). SP: Participants began with elbows fully extended, descended to ~90° of elbow flexion at a controlled, self-selected tempo, and then performed a concentric push to full extension (Fan et al., 2025). PP: From full elbow extension, participants executed a rapid countermovement to ~90° of elbow flexion, followed by an explosive concentric push that propelled the body off the force plates (Dhahbi et al., 2017b; Zalleg et al., 2020). Upon landing, participants minimized impact and returned to the starting position in a controlled manner (Dhahbi et al., 2017b). SSP: Participants began from a static bottom position with elbows flexed to ~90° and performed a maximal-effort concentric action to lift the body off the plates, without a preceding stretch preloading phase (Wang et al., 2017; Zalleg et al., 2020). Each participant completed three valid trials for each task. A 1-min rest interval was provided between consecutive trials, and a 3-min rest interval was provided between task conditions. All trials were visually inspected by experienced researchers to ensure proper task execution. Motion-capture data were collected at 120 Hz using a three-dimensional motion capture system with 13 cameras (Motive 2.2.0; OptiTrack, NaturalPoint Inc., Corvallis, OR, USA). Two reflective markers were placed on the bilateral acromion, and the motion-capture data were used solely for event detection and analysis-window definition, not for joint-angle analysis (Dhahbi et al., 2017a; Fan et al., 2025). Surface EMG signals from 12 right-upper-limb muscles were recorded at 1200 Hz using wireless Trigno Avanti sensors (Delsys Inc., Natick, MA, USA). Sensors were positioned over the following muscles according to Atlas of Muscle Innervation Zones (Barbero et al., 2012) with electrodes aligned parallel to the presumed muscle-fiber direction. Specifically, sensors were placed over flexor carpi radialis (FCR) at 37-100% of the medial epicondyle-radial styloid anatomical landmark frame (ALF); extensor carpi radialis (ECR) at 42-100% of the styloid-process midpoint-lateral supracondylar ridge ALF; biceps brachii (BB) before 62% of the acromion-distal biceps tendon ALF; triceps brachii (TB) at 66-100% of the acromial angle-epicondyle ALF; |process-deltoid tuberosity ALF; posterior deltoid (PD) perpendicular to the acromion-glenoid line at approximately 47%; pectoralis major (PM) at 76-100% of the acromial angle-xiphoid process ALF; serratus anterior (SA) between the fifth and seventh ribs; upper trapezius (UT) at 0-44% of the distal clavicle-C6 spinous process ALF; latissimus dorsi (LD) at 55-100% of the posterior superior iliac spine-upper humerus ALF; erector spinae (ES) lateral to the T12-L5 spinous-process line; and rectus abdominis (RA) over the right abdominal muscle belly, parallel to the linea alba. Prior to sensors placement, the skin was shaved and cleansed with alcohol to reduce impedance (Fan et al., 2025), after which all sensors were secured using elastic adhesive tape. GRF were recorded synchronously at 1200 Hz using OR6-6-2000 force plates (Advanced Mechanical Technology Inc., Watertown, MA, USA). Motion-capture, EMG, and force-plate data were processed in Visual3D (C-Motion Inc., Germantown, MD, USA) for three-dimensional reconstruction, event identification, and definition of analysis windows. The lowest vertical acromion position was defined as the onset of the concentric phase. The endpoint of SP was defined as peak acromion height, whereas the endpoints of PP and SSP were defined as take-off to reflect their explosive concentric nature. These endpoints were selected to represent the task-specific completion of the concentric propulsion phase. For SP, peak acromion height reflected the end of the upward pushing action, whereas take-off represented the completion of the explosive concentric phase in PP and SSP. Using these boundaries, EMG and vertical GRF signals were extracted and processed in MATLAB R2021a (MathWorks, Natick, MA, USA). GRF signals were normalized to body mass (N/kg), and time-normalized to 101 points. EMG signals were processed with band-pass filtering (20-450 Hz; 4th-order Butterworth), full-wave rectification, and low-pass filtering (6 Hz; 4th-order Butterworth) to obtain linear envelopes (Fan et al., 2024). Each muscle’s EMG envelope was normalized to the peak activation of the same muscle across all valid trials of the three pushing tasks (Fan et al., 2025), and then stored as a 12 × 101 EMG matrix (12 muscles × 101 time points). For each participant and each task, the two trials showing the highest within-task similarity among the three valid trials were retained, based on the intraclass correlation coefficient calculated from the time-normalized EMG envelopes across the 12 recorded muscles (Fan et al., 2024). The retained two trials were then averaged to obtain a representative EMG matrix for synergy extraction. Processed EMG matrices were analyzed using non-negative matrix factorization (NNMF), a widely used method for identifying low-dimensional neuromuscular control strategies (Fan et al., 2024; 2025). For each trial, NNMF decomposed the EMG matrix into a muscle weighting matrix (M; synergy modules) and a temporal activation matrix (P; synergy primitives):
M is a 12 × x matrix and P is an x × 101 matrix for the identified synergies (x is the number of synergies). The error term represents the residual between original and reconstructed EMG. NNMF was repeated 20 times to avoid local minima, and the solution with the lowest reconstruction error was selected for each pushing condition (Fan et al., 2024). The optimal number of synergies (x) was determined using global variance accounted for (VAF), with VAF > 0.95 indicating acceptable reconstruction (Fan et al., 2025; Li et al., 2022). VAF was computed as:
To enable cross-task comparison, the number of synergies was fixed at k = 2 for all conditions. Synergy primitives were classified using k-means clustering (Fan et al., 2024), repeated 50 times with different random seeds to reduce stochastic variability (Fan et al., 2025). Cluster quality was evaluated using silhouette scores, and the solution with the highest score was selected. The resulting clustered synergy primitives and their corresponding modules were used for further analysis.
Statistical analysisStatistical analyses were conducted to examine task-specific differences among the three pushing conditions. Cosine similarity (CS) was first computed to quantify the similarity of synergy modules between tasks, with values closer to 1 indicating greater similarity (Santuz et al., 2017). Differences in individual muscle weights within each synergy module were evaluated using paired-sample t-tests, with normality assessed prior to testing; when normality was violated, Wilcoxon signed-rank tests were used. The Benjamini-Hochberg false discovery rate procedure was applied for multiple-comparison correction. Effect sizes for discrete paired comparisons were expressed as bias-corrected Hedges’ g for within-subject comparisons to reduce small-sample bias, with 95% confidence intervals calculated using the same correction procedure. One-dimensional Statistical Parametric Mapping (SPM1d) with a one-way ANOVA model (SPM{F}) was used to assess task-specific differences in synergy primitives and force-time profiles. Significant main effects (P < 0.05) were followed by paired SPM{t} post-hoc tests with a Bonferroni-adjusted α level of 0.017 (0.05/3) (Fan et al., 2024; Hughes et al., 2022). Parametric SPM1d analyses were used for the primary trajectory comparisons, and the normality assumption of the trajectory residuals was assumed for these analyses. SPM{F} and SPM{t} results were reported with suprathreshold time intervals and cluster-level P values (Pataky et al., 2015). For significant post-hoc trajectory differences, pointwise paired standardized mean differences were calculated at each normalized time point, corrected for small-sample bias, and averaged within each suprathreshold cluster. These values were reported as mean cluster Hedges’ g. All analyses were performed in MATLAB R2021a, with statistical significance set at α < 0.05 unless otherwise specified.
Muscle synergy extractionNeuromuscular control across the three pushing tasks was consistently characterized by two muscle synergies (SP VAF = 0.980 ± 0.008, PP VAF = 0.966 ± 0.013, SSP VAF = 0.967 ± 0.011; VAF > 0.95). The VAF curves showed a plateau after two synergies across the three tasks (Figure 1). Accordingly, each task was decomposed into two synergy modules and two synergy primitives. Synergy primitive 1 was primarily active during the early concentric phase (0-50%), whereas synergy primitive 2 peaked during the later phase (50-100%) (Figure 2).
Synergy module characteristicsFor synergy modules, synergy module 1 showed consistently low cosine similarity across all task pairs (SP vs. PP: CS = 0.78; SP vs. SSP: CS = 0.79; PP vs. SSP: CS = 0.76), all of which were below the commonly accepted similarity threshold (< 0.90). Synergy module 2 exhibited a similar pattern, with low similarity between SP vs. PP (CS = 0.83) and SP vs. SSP (CS = 0.81), whereas PP vs. SSP showed moderately higher similarity (CS = 0.88), though still below the 0.90 criterion (Figure 3). Paired comparisons revealed that, in synergy module 1, PP showed lower FCR (P = 0.001, Hedges’ g = -1.008, 95% CI [-2.135, -0.463]) and UT (P < 0.001, Hedges’ g = -1.302, 95% CI [-2.013, -0.946]), but higher RA contributions (P < 0.001, Hedges’ g = 1.156, 95% CI [0.545, 2.398]) compared with SP. SSP also showed lower FCR weighting compared with SP (P < 0.001, Hedges’ g = -1.114, 95% CI [-1.893, -0.692]). Compared with SSP, PP showed higher RA contribution (P = 0.002, Hedges’ g = 0.931, 95% CI [0.736, 1.330]). No other muscle-weight differences in synergy module 1 remained significant after correction. In synergy module 2, no significant differences were observed between PP and SSP. Compared with SP, PP showed higher FCR (P < 0.001, Hedges’ g = 1.518, 95% CI [0.937, 2.969]), SA (P = 0.002, Hedges’ g = 0.907, 95% CI [0.396, 2.150]), and UT contributions (P < 0.001, Hedges’ g = 1.312, 95% CI [0.792, 2.291]). SSP showed higher FCR (P < 0.001, Hedges’ g = 1.536, 95% CI [1.124, 2.386]), TB (P = 0.003, Hedges’ g = 0.877, 95% CI [0.316, 2.052]), and UT contributions (P = 0.006, Hedges’ g = 0.787, 95% CI [0.349, 1.490]) compared with SP. No other muscle-weight differences in synergy module 2 remained significant after correction (Figure 4).
Synergy primitive characteristicsSPM analysis revealed significant task-specific differences in synergy primitive 1 during 0-12% (df = 2, 28; F* = 7.299, P = 0.030) and 48-71% (P < 0.001) of the movement cycle. Post-hoc comparisons indicated that SSP demonstrated significantly higher activation than SP during 49-68% of the cycle (df = 14; t* = 4.312, P < 0.001; mean cluster Hedges’ g = 1.42). Conversely, SSP showed significantly lower activation than PP during 0-5% of the cycle (df = 14; t* = 4.983, P = 0.006; mean cluster Hedges’ g = -1.31). No significant difference was observed between SP and PP. For synergy primitive 2, SPM analysis identified significant task-specific differences during 71-100% of the movement cycle (df = 2, 28; F* = 7.131, P < 0.001). Post-hoc analyses showed that PP exhibited significantly higher activation than SP during 71-100% of the cycle (df = 14; t* = 4.239, P < 0.001; mean cluster Hedges’ g = 1.06), and SSP exhibited significantly higher activation than SP during 80-100% of the cycle (df = 14; t* = 4.254, P < 0.001; mean cluster Hedges’ g = 1.29). No significant difference was observed between PP and SSP (Figure 5).
Force-Time Profile CharacteristicsBefore time normalization, the absolute duration of the concentric phase was 0.80 ± 0.08 s for SP, 0.58 ± 0.05 s for PP, and 0.67 ± 0.08 s for SSP. SPM analysis identified significant task-specific differences in vertical force during 0-16% (df = 2, 28; F* = 6.308, P = 0.012), 24-85% (P < 0.001), and 90-100% (P = 0.033) of the concentric phase. Post-hoc analyses showed that PP produced significantly higher force than SP during 13-79% of the cycle (df = 14; t* = 4.038, P < 0.001; mean cluster Hedges’ g = 2.63), but significantly lower force during 92-100% of the cycle (P = 0.009; mean cluster Hedges’ g = -1.53). SSP generated significantly lower force than SP during 0-12% (df = 14; t* = 3.810, P = 0.009; mean cluster Hedges’ g = -1.69) and 90-100% (P = 0.015; mean cluster Hedges’ g = -1.12), but significantly higher force during 33-36% (P = 0.016; mean cluster Hedges’ g = 0.94) and 56-83% (P < 0.001; mean cluster Hedges’ g = 1.79). Additionally, SSP produced significantly lower force than PP during 0-14% of the cycle (df = 14; t* = 4.009, P = 0.001; mean cluster Hedges’ g = -2.24), but significantly higher force during 79-80% of the cycle (P = 0.016; mean cluster Hedges’ g = 0.98) (Figure 5).
This study compared the neuromuscular control strategies and force-time profiles of three upper-limb pushing tasks (SP, PP, and SSP) during the concentric phase. Two key findings emerged. First, although all tasks were governed by two primary muscle synergies, both synergy structures and temporal primitives exhibited clear task-specific differences, indicating distinct neuromuscular coordination strategies. Second, PP and SSP generated markedly higher force during the mid-to-late concentric phase compared with SP, accompanied by stronger late-phase synergy activation. Collectively, these findings support our hypotheses and demonstrate that, despite sharing similar gross movement patterns, the three pushing tasks exhibit distinct neuromechanical coupling during force generation. These results suggest that intermuscular coordination patterns vary according to differing mechanical demands. Building on these findings, the observation that two synergies were sufficient to reconstruct all tasks suggests a relatively low-dimensional organization of neuromuscular control during the concentric phase (Eken et al., 2023). Synergy module 1 peaked during the early concentric phase, whereas synergy module 2 peaked during the latter half, reflecting a sequential organization of neuromuscular activation that is characteristic of upper-limb pushing actions. Notably, despite the conserved number of synergies, the three tasks exhibited clear task-specific differences in both synergy structure and activation timing. This finding suggests that performing similar movement patterns under different force demands is associated with changes in muscle-weighting distributions within each synergy (Fan et al., 2024), rather than simply scaling the activation magnitude of individual muscles. These observations are consistent with, yet extend, previous research on pushing tasks (Dhahbi et al., 2017b; Fan et al., 2025; Koch et al., 2012; Nichols and Szivak, 2021; Zalleg et al., 2020), by demonstrating that task-specific adjustments occur not only in activation magnitude but also in the underlying synergy organization. Earlier studies manipulating hand position or support width consistently showed an invariant number of synergies across push-up variations (Fan et al., 2025). Prior studies have also indicated that kneeling PP elicit higher activation in the PM, TB, and AD compared with traditional PP (García-Massó et al., 2011). Moreover, alterations in task intensity or load requirements have been shown to modify the contributions of major muscles involved in upper-limb propulsion. Building upon this body of evidence, our results further show that although the number of synergies remained constant across SP, PP, and SSP, both the internal weighting structure and the temporal activation profiles exhibited clear and systematic task specificity. This suggests that, under distinct mechanical demands, the observed EMG-derived synergy patterns do not simply reflect the scaling of a fixed synergy structure; rather, they indicate task-dependent adjustments in synergy composition and activation timing. In synergy module 1, both PP and SSP exhibited lower weights for FCR and UT, but higher weights for RA and SA. This pattern suggests an increased demand for trunk stabilization (Saeterbakken et al., 2022) and scapular control (Umehara et al., 2018) during the initiation of explosive pushing actions (García-Massó et al., 2011). In contrast, the higher FCR and UT weights observed in the SP reflect its greater emphasis on forearm-mediated motor control (Wilke and Krause, 2019). These task-specific differences suggest that, under higher-velocity or higher-force demands, the observed EMG-derived synergy patterns may reflect reduced contributions from distal fine-control muscles and greater involvement of scapular-trunk stabilizers during proximal-to-distal force transmission. This interpretation is consistent with the findings of Krings et al.,(Krings et al., 2021), who reported decreased upper-arm muscle activity but increased shoulder muscle activation under higher loading demands. Additionally, synergy module 2 demonstrated greater weighting of FCR, TB, UT, and SA in both PP and SSP. This pattern indicates the critical role of scapular and trunk stabilization during the terminal phase of high-speed or high-load pushing (Saeterbakken et al., 2022), and reflecting the importance of wrist-forearm stability in maintaining distal rigidity to minimize energy loss in the kinetic chain. The increased contribution of TB, the primary elbow extensor (Kholinne et al., 2018), further suggests that forceful elbow extension is essential for achieving peak concentric output, particularly in high-intensity tasks. However, the lack of differences in AD, PD, and PM suggests that the observed pattern may reflect a relatively consistent proximal stabilization strategy across tasks (Bagherian et al., 2019). The temporal organization of the synergy primitives further revealed distinct task-specific patterns of synergy activation. Differences in synergy primitive 1 during the early concentric phase indicate that PP elicited greater neuromuscular activation at movement onset, likely reflecting feedforward-like muscle excitation and SSC-related pre-activation during the rapid eccentric-concentric transition (García-Massó et al., 2011). This mechanism may also account for the elevated early-phase force observed in PP. This interpretation aligns with Dhahbi et al., (2017b), who similarly reported superior early-phase force production during PP actions. Moreover, SSP showed significantly higher activation than SP during the late-early to mid-portion of synergy primitive 1 (49-68% phase), and PP demonstrated a comparable upward trend. In synergy primitive 2, both PP and SSP demonstrated consistently higher activation than SP across a broad portion of the late concentric phase. This suggests that these tasks require greater propulsive capacity and enhanced stabilization to accelerate the body or achieve lift-off (García-Massó et al., 2011). This neuromechanical pattern corresponds closely with the elevated mid-to-late concentric forces observed during the mid-to-late phase (24-85% phase). Taken together, these findings indicate that mechanical performance in upper-limb pushing tasks is governed by the phase-dependent organization of muscle synergies. This reinforces the value of synergy analysis as a sensitive approach for uncovering the underlying neuromechanical strategies that shape task-specific force production. SSP also exhibited greater force than PP during the terminal phase of the concentric action (79-80%), suggesting subtle task-specific differences in force expression near the end of propulsion. Hughes et al.,(Hughes et al., 2022) demonstrated that SPM is highly sensitive for detecting changes across the full force-time profile. Additionally, differences in force-velocity profiles under varying loads (Samozino et al., 2018) indicate that PP should exhibit superior end-phase velocity and longer flight time compared with SSP (Dhahbi et al., 2017b), which is consistent with our findings. Notably, PP and SSP did not differ in synergy primitive 2 or in the structural organization of synergy module 2. This suggests that, despite differences in mechanical output, both tasks rely on a comparable underlying synergy structure and muscle-coordination strategy during the terminal portion of the concentric phase. This finding highlights that variations in mechanical output do not necessarily reflect changes in motor-control strategies, thereby underscoring the distinct methodological value of synergy analysis for elucidating the neuromuscular control patterns underlying movement regulation. This study has several limitations that should be acknowledged. First, the sample consisted exclusively of well-trained male rugby athletes, which restricts the generalizability of the findings; future studies should recruit female athletes, participants from other sports, and non-athlete populations to determine whether the observed neuromuscular control patterns are broadly applicable. In addition, although the a priori sample-size justification supported the inclusion of 15 participants for a scalar repeated-measures ANOVA, the power characteristics of SPM1D trajectory analyses may differ; therefore, the present sample size should be interpreted as a conservative lower-bound estimate for the trajectory-based analyses. Second, only 12 upper-limb and trunk muscles were recorded due to constraints in EMG sensor availability, which may have influenced the dimensionality and interpretation of the extracted synergies (Fan et al., 2024). Third, because of limitations in data acquisition, particularly the inability to capture the eccentric phase in SSP, the analysis was restricted to the concentric phase, limiting insight into eccentric-concentric and SSC-related interactions. Future research should therefore incorporate a broader set of muscles and include both eccentric and concentric phases to more comprehensively characterize how the SSC modulates synergy structure and neuromuscular coordination. Practical ApplicationsThe findings of this study provide practical implications for exercise selection and training design aimed at addressing different upper-limb pushing demands. The PP was characterized by greater early-to-mid phase force production and stronger synergy activation, making it a relevant option when the goal is to emphasize explosive upper-body pushing characteristics and rate-dependent force expression. In contrast, the SSP produced sustained and elevated force output during the late concentric phase, suggesting that it may be useful for targeting end-range force production and proximal stability demands. The increased weighting of SA, UT, and RA underscores the important role of the scapular-trunk complex in high-force pushing tasks. Conversely, the elevated contributions of FCR and TB highlight the potential importance of distal joint stability and elbow-extension capacity for efficient force transmission along the upper-limb kinetic chain. Accordingly, coaches and practitioners can use these task-specific neuromechanical characteristics as a reference when selecting push-up variations for different upper-limb pushing demands.
This study shows that although the three pushing tasks share the same number of muscle synergies, their synergy structures, temporal activation profiles, and concentric force-time characteristics differ markedly. These findings indicate that upper-limb pushing tasks with different mechanical demands are associated with distinct intermuscular-coordination patterns. PPs are characterized by rapid force development and prominent early-phase neuromuscular activation, whereas SSPs show more sustained synergy activation throughout the concentric phase. Collectively, these findings advance understanding of the neuromuscular control mechanisms underpinning upper-limb pushing actions and may inform exercise selection for athletic populations with similar upper-limb pushing demands.
| ACKNOWLEDGEMENTS |
This study was funded by the Scientific Research Project of the College of Education and Sports Sciences, Yangtze University (JTKZ01), and the Undergraduate Innovation and Entrepreneurship Training Program of Yangtze University (Yz2025186). The authors report no actual or potential conflicts of interest. While the datasets generated and analyzed in this study are not publicly available, they can be obtained from the corresponding author upon request. All experimental procedures were conducted in compliance with the relevant legal and ethical standards of the country where the study was carried out. The authors declare that no Generative AI or AI-assisted technologies were used in the writing of this manuscript. |
|
| AUTHOR BIOGRAPHY |
|
 |
Penglei Fan |
| Employment: College of Education and Sports Sciences, Yangtze University, Jingzhou, Hubei, China |
| Degree: PhD |
| Research interests: Sports Biomechanics, Neuromuscular Control Strategies, Sports training |
| E-mail: fanpenglei@yangtzeu.edu.cn |
| |
 |
Mengzhao Han |
| Employment: College of Education and Sports Sciences, Yangtze University, Jingzhou, Hubei, China |
| Degree: MS |
| Research interests: Sports Biomechanics, Sports training |
| E-mail: hanmengzhao.stu@yangtzeu.edu.cn |
| |
 |
Ting Wang |
| Employment: Institute of Student Physical Health Promotion, Jingchu University of Technology, Jingmen, Hubei, China |
| Degree: PhD |
| Research interests: Sports Biomechanics, Sports injury prevention, athletic performance |
| E-mail: WangTing@jcut.edu.cn |
| |
 |
Guihua Huang |
| Employment: College of Education and Sports Sciences, Yangtze University, Jingzhou Hubei, China |
| Degree: MS |
| Research interests: Sport and Exercise Biomechanics, Sports training |
| E-mail: 2189711766@qq.com |
| |
 |
Xiaoshuai Wang |
| Employment: College of Education and Sports Sciences, Yangtze University, Jingzhou Hubei, China |
| Degree: MS |
| Research interests: Sport and Exercise Biomechanics, Sports training |
| E-mail: 13317215136@163.com |
| |
 |
Ming Li |
| Employment: College of Education and Sports Sciences, Yangtze University, Jingzhou Hubei, China |
| Degree: PhD |
| Research interests: Sports Biomechanics, strength and conditioning, athletic performance |
| E-mail: lming@yangtzeu.edu.cn |
| |
|
| |
| REFERENCES |
 Bagherian, S., Ghasempoor, K., Rahnama, N., Wikstrom, E.A. (2019) The effect of core stability training on functional movement patterns in college athletes. Journal of Sport Rehabilitation 28, 444-449. Crossref |
Baiget, E., Colomar, J., Corbi, F. (2021) Upper-limb force-time characteristics determine serve velocity in competition tennis players. International Journal of Sports Physiology and Performance 17, 358-366. Crossref |
Barbero, M., Merletti, R., Rainoldi, A. (2012) Atlas of Muscle Innervation Zones: Understanding Surface Electromyography and Its Applications. Springer Science & Business Media. Crossref |
Del Vecchio, A. (2023) Neuromechanics of the rate of force development. Exercise and Sport Sciences Reviews 51, 34. Crossref |
Dhahbi, W., Chaabene, H., Chaouachi, A., Padulo, J., Behm, D.G., Cochrane, J., Burnett, A., Chamari, K. (2022) Kinetic analysis of push-up exercises: A systematic review with practical recommendations. Sports Biomechanics 21, 1-40. Crossref |
Dhahbi, W., Chaouachi, A., Cochrane, J., Chèze, L., Chamari, K. (2017a) (2017a) Methodological issues associated with the use of force plates when assessing push-ups power. Journal of Strength and Conditioning Research 31, e74. Crossref |
Dhahbi, W., Chaouachi, A., Dhahbi, A.B., Cochrane, J., Chèze, L., Burnett, A., Chamari, K. (2017b) (2017b) The effect of variation of plyometric push-ups on force-application kinetics and perception of intensity. International Journal of Sports Physiology and Performance 12, 190-197. Crossref |
Eken, M.M., Arnold, S.L., Thijssen, J., van der Schaaf, M., Engelbrecht, L., Lamberts, R.P. (2023) The influence of a multistage mountain-bike race on neuromuscular activation and synergies: A case study. International Journal of Sports Physiology and Performance 19, 315-321. Crossref |
Fan, P., Kim, Y., Tang, X., Wang, T., Li, B., Kim, S. (2025) Effects of hand positions on neuromuscular control strategies in closed-kinetic-chain push-ups. Journal of Men’s Health 21, 30-38. Crossref |
Fan, P., Yang, Z., Wang, T., Li, J., Kim, Y., Kim, S. (2024) Neuromuscular control strategies in basketball shooting: Distance-dependent analysis of muscle synergies. Journal of Sports Science and Medicine 23, 571-580. Crossref |
Frère, J., Hug, F. (2012) Between-subject variability of muscle synergies during a complex motor skill. Frontiers in Computational Neuroscience 6. Crossref |
García-Massó, X., Colado, J.C., González, L.M., Salvá, P., Alves, J., Tella, V., Triplett, N.T. (2011) Myoelectric activation and kinetics of different plyometric push-up exercises. Journal of Strength and Conditioning Research 25, 2040. Crossref |
Hughes, S., Warmenhoven, J., Haff, G.G., Chapman, D.W., Nimphius, S. (2022) Countermovement jump and squat jump force-time curve analysis in control and fatigue conditions. Journal of Strength and Conditioning Research 36, 2752. Crossref |
Kholinne, E., Zulkarnain, R.F., Sun, Y.C., Lim, S., Chun, J.-M., Jeon, I.-H. (2018) The different role of each head of the triceps brachii muscle in elbow extension. Acta Orthopaedica et Traumatologica Turcica 52, 201-205. |
Koch, J., Riemann, B.L., Davies, G.J. (2012) Ground reaction force patterns in plyometric push-ups. Journal of Strength and Conditioning Research 26, 2220. Crossref |
Krings, B.M., Shepherd, B.D., Swain, J.C., Turner, A.J., Chander, H., Waldman, H.S., McAllister, M.J., Knight, A.C., Smith, J.W. (2021) Impact of fat grip attachments on muscular strength and neuromuscular activation during resistance exercise. Journal of Strength and Conditioning Research 35, S152. Crossref |
Li, Z., Zhao, X., Wang, Z., Xu, R., Meng, L., Ming, D. (2022) A hierarchical classification of gestures under two force levels based on muscle synergy. Biomedical Signal Processing and Control 77, 103695. Crossref |
Lum, D., Soh, S.K., Teo, C.J.H., Wong, O.Q.H., Lee, M.J.C. (2022) Effects of performing isometric bench press training at single versus multiple joint positions on strength and power performance. International Journal of Sports Physiology and Performance 17, 1061-1069. Crossref |
McKay, A.K.A., Stellingwerff, T., Smith, E.S., Martin, D.T., Mujika, I., Goosey-Tolfrey, V.L., Sheppard, J., Burke, L.M. (2021) Defining training and performance caliber: A participant classification framework. Crossref |
Nichols, I.A., Szivak, T.K. (2021) Effects of different hand widths on plyometric push-up performance. Journal of Strength and Conditioning Research 35, S80. Crossref |
Nicol, C., Avela, J., Komi, P.V. (2006) The stretch-shortening cycle: A model to study naturally occurring neuromuscular fatigue. Sports Medicine 36, 977-999. Crossref |
Ning, C., Sheykhlouvand, M. (2025) Selected immunoendocrine and physiological performance adaptations to different volume of upper-body plyometric training in national-level male volleyball players. International Journal of Sports Physiology and Performance 20, 363-371. Crossref |
Pataky, T.C., Vanrenterghem, J., Robinson, M.A. (2015) Zero- vs. one-dimensional, parametric vs. non-parametric, and confidence interval vs. hypothesis testing procedures in one-dimensional biomechanical trajectory analysis. Journal of Biomechanics 48, 1277-1285. Crossref |
Rogers, D.K., McKeown, I., Parfitt, G., Burgess, D., Eston, R.G. (2020) Effect of biological maturation on performance of the athletic ability assessment in Australian rules football players. International Journal of Sports Physiology and Performance 16, 28-36. Crossref |
Saeterbakken, A.H., Stien, N., Pedersen, H., Andersen, V. (2022) Core muscle activation in three lower extremity exercises with different stability requirements. Journal of Strength and Conditioning Research 36, 304. Crossref |
Samozino, P., Rivière, J.R., Rossi, J., Morin, J.-B., Jimenez-Reyes, P. (2018) How fast is a horizontal squat jump? International Journal of Sports Physiology and Performance 13, 910-916. Crossref |
Santuz, A., Ekizos, A., Janshen, L., Baltzopoulos, V., Arampatzis, A. (2017) On the methodological implications of extracting muscle synergies from human locomotion. International Journal of Neural Systems 27, 1750007. Crossref |
Speranza, M.J.A., Gabbett, T.J., Greene, D.A., Johnston, R.D., Townshend, A.D., O’Farrell, B. (2018) An alternative test of tackling ability in rugby league players. International Journal of Sports Physiology and Performance 13, 347-352. Crossref |
Umehara, J., Kusano, K., Nakamura, M., Morishita, K., Nishishita, S., Tanaka, H., Shimizu, I., Ichihashi, N. (2018) Scapular kinematic and shoulder muscle activity alterations after serratus anterior muscle fatigue. Journal of Shoulder and Elbow Surgery 27, 1205-1213. Crossref |
Wang, R., Hoffman, J.R., Sadres, E., Bartolomei, S., Muddle, T.W.D., Fukuda, D.H., Stout, J.R. (2017) Evaluating upper-body strength and power from a single test: The ballistic push-up. Journal of Strength and Conditioning Research 31, 1338. Crossref |
Wilke, J., Krause, F. (2019) Myofascial chains of the upper limb: A systematic review of anatomical studies. Crossref |
Zalleg, D., Ben Dhahbi, A., Dhahbi, W., Sellami, M., Padulo, J., Souaifi, M., Bešlija, T., Chamari, K. (2020) Explosive push-ups: From popular simple exercises to valid tests for upper-body power. Journal of Strength and Conditioning Research 34, 2877. Crossref |
|
| |
|
|
|
|