|
|
|
| |
| ABSTRACT |
|
The purpose of this study was to determine whether the excessive spikes observed in the surface electromyography (sEMG) spectrum recorded during whole-body vibration (WBV) exercises contain motion artifacts and/or reflex activity. The occurrence of motion artifacts was tested by electrical recordings of the patella. The involvement of reflex activity was investigated by analyzing the magnitude of the isolated spikes during changes in voluntary background muscle activity. Eighteen physically active volunteers performed static squats while the sEMG was measured of five lower limb muscles during vertical WBV using no load and an additional load of 33 kg. In order to record motion artifacts during WBV, a pair of electrodes was positioned on the patella with several layers of tape between skin and electrodes. Spectral analysis of the patella signal revealed recordings of motion artifacts as high peaks at the vibration frequency (fundamental) and marginal peaks at the multiple harmonics were observed. For the sEMG recordings, the root mean square of the spikes increased with increasing additional loads (p < 0.05), and was significantly correlated to the sEMG signal without the spikes of the respective muscle (r range: 0.54 - 0.92, p < 0.05). This finding indicates that reflex activity might be contained in the isolated spikes, as identical behavior has been found for stretch reflex responses evoked during direct vibration. In conclusion, the spikes visible in the sEMG spectrum during WBV exercises contain motion artifacts and possibly reflex activity. |
| Key words:
Stretch reflex, vibration training, power spectral density, frequency analysis, filtering, spectral linear interpolation
|
Key
Points
- The spikes observed in the sEMG spectrum during WBV exercises contain motion artifacts and possibly reflex activity
- The motion artifacts are more pronounced in the first spike than the following spikes in the sEMG spectrum
- Reflex activity during WBV exercises is enhanced with an additional load of approximately 50% of the body mass
|
Whole-body vibration (WBV) exercises acutely increase muscle activity during the exposure, as shown in numerous studies with the help of surface electromyography (sEMG) recordings (Cardinale and Lim, 2003; Di Giminiani et al., 2013; Krol et al., 2011; Marín et al., 2012; Perchthaler et al., 2013). While the exact underlying mechanisms remain unclear, reflex muscular contractions have been proposed (Cardinale and Bosco, 2003). During directly applied vibration, the primary endings of muscle spindles respond up to frequencies of 180 Hz (Roll and Vedel, 1982), whereas secondary endings are less sensitive to vibration and respond at lower frequencies (Brown et al., 1967; Burke et al., 1976a). In addition to the spindle afferents, golgi tendon organs are responsive to vibration (Burke et al., 1976a). Besides reflex activity, motion artifacts are assumed to occur, visible in the sEMG spectrum as excessive spikes at the vibration frequency and its multiple harmonics (Abercromby et al., 2007). As the reflex activity (Ritzmann et al., 2010) and the motion artifacts (Fratini et al., 2009) are both phase-locked to the vibration frequency, their influence on sEMG signals is diffficult to predict. Occurrence of motion artifacts during WBV exercises has been evidenced by signal recordings with so-called dummy electrodes. Electrodes have been placed on the patella assuming that the recorded signal would be related to motion artifacts only (Fratini et al., 2009). Although excessive peaks in the sEMG spectrum were located, it cannot be excluded that these peaks were not sEMG recordings from surrounding muscles. To address this limitation, a similar set-up has been repeated (Ritzmann et al., 2010; Sebik et al., 2013) by positioning electrodes on the muscle with several layers of insulating tape between skin and electrodes. While Sebik et al. (Sebik et al., 2013) found motion artifact-contaminated sEMG signals, Ritzmann et al. (Ritzmann et al., 2010) observed only a marginal contribution. As a consequence, there is no consensus whether the spikes in the sEMG spectrum are contaminated by motion artifacts. The most frequently cited mechanism by which WBV increases muscle activity is the tonic vibration reflex (TVR) (Cardinale and Bosco, 2003; Torvinen et al., 2002). The TVR has been well proven for locally applied vibration on the muscle belly or the tendon (Burke et al., 1976b; Eklund and Hagbarth, 1966), but with no conclusive evidence that the TVR is applicable to WBV. However, almost identical latencies between actual stretch reflexes and WBV-induced reflex responses have been reported (Ritzmann et al., 2010). Further, a study by Pollock et al. (2012) has shown a strong relation between the timing of motor unit firings and the phase of the vibration cycle using intramuscular sEMG. Additionally, motor unit synchronization has been shown to take place during WBV by means of multi motor unit recordings and sEMG signal rectification (Sebik et al., 2013). Interestingly, stretch reflex responses evoked via directly-applied vibration are enhanced by muscle contractions (Burke et al., 1976b; Eklund and Hagbarth, 1966). If WBV elicits stretch reflexes, the magnitude of the spikes in the sEMG spectrum is expected to increase with increasing background muscle activity. Surprisingly, this hypothesis has never been examined. Therefore, the goal of this study was to determine whether sEMG signals during WBV exercises contain motion artifacts and/or reflex activity. The latter was tested by analyzing the magnitude of the isolated spikes in the sEMG power spectrum using different background muscle activity levels. Contribution of motion artifacts were verified with the help of electrical recordings from the patella during the vibration. We hypothesized that (1) the sEMG measured during WBV contains motion artifacts caused by the vibration of the platform, that (2) motion artifacts are more prevalent in the first spike than in the following spikes, and that (3) the magnitude of the spikes in the sEMG spectrum increases with increasing voluntary background muscle activity. ParticipantsEighteen (10 men, 8 women) physically active students (mean ± SD, age: 23.8 ± 3.2 years; height: 1.73 ± 0.08 m; body mass: 67.8 ± 10.9 kg) volunteered to participate in the study. The exclusion criteria included frequent practice of WBV and/or neurological injuries to the lower extremity. The study was conducted according to the Helsinki Declaration (1964) and the study protocol was approved by the local ethics committee. Participants were informed about potential risks and signed written consent forms prior to their participation.
Experimental designIn order to evaluate if the sEMG signals during WBV contain motion artifacts (hypothesis 1) and if they are more pronounced in the first spike than the following spikes (hypothesis 2), electrical signals from the patella were collected during the vibration. To test hypothesis (3), i.e., the magnitude of the spikes in the sEMG spectrum increases with increasing voluntary background muscle activity, sEMG signals of five lower limb muscles were recorded during static squat exercises with and without WBV using no load and an additional load of 33 kg. The additional load was used to increase the voluntary background muscle activity. The WBV device used in this study was a synchronous oscillating platform (Power Plate pro6, North America, Northbrook, IL, USA), which delivers vibration in the vertical direction. The vibration frequency was set at 30 Hz, and the peak-to-peak displacement of the platform was 2 mm (preliminarily evaluated via double integration of the platform acceleration signal). The experimental session consisted of four vibration conditions presented in a randomized order, including two WBV trials (no load, 33 kg load) and two control trials without vibration (no load, 33 kg load). WBV trials consisted of 20-s static squats with a 1-min break in between trials. All participants were familiarized with WBV previously to the test session. The additional load was applied via a standard weightlifting bar measuring 150 cm in length with a weight of 5 kg; weight plates were attached to each side to obtain the 33 kg load. For each trial, the bar was positioned on the shoulders of the subjects, and all participants were familiar with loaded squat techniques. The participants were barefoot during the vibration trials in order to avoid damping effects due to different footwear, and foot position was marked on the platform in order to have similar stance between subjects. The participants were advised to flex their knees at an angle of 70° (0° corresponding to full extension) during the WBV exercises, which was monitored and recorded with the help of an electrical twin-axis goniometer (TSD130B, Biopac® Systems Inc., Holliston, MA, USA). When a participant could not maintain the position throughout the exercise, trials were repeated. Subsequent analysis showed that the participants performed their squats at a knee angle of 68.2 ± 1.3° (mean ± SD). The acceleration of the vibration stimuli was controlled and recorded with the help of a tri-axial accelerometer (50G, TSD109F, Biopac® Systems Inc., Holliston, MA, USA) that was placed on the platform in alignment to the third toe (Lorenzen et al., 2009). Root mean square (RMS) calculation of the vertical platform acceleration proved a RMS (mean ± SD) of 2.4 ± 0.1 g, with identical values between loads.
sEMG and patella signal recordingPairs of bipolar silver-chloride electrodes (10-mm diameter, Contrôle Graphique Medical, Brie-Comte-Robert, France) were positioned over the muscle bellies of the vastus lateralis (VL), rectus femoris (RF), biceps femoris (BF), soleus (SOL), and gastrocnemius lateralis (GL) of the right lower limb to record sEMG activity. The longitudinal axes of the electrodes were placed in line with the presumed direction of the respective underlying muscle fibers, with a center-to-center distance of 20 mm. The reference electrode was attached to the left patella. Low interelectrode resistance (< 5 k™¦) was obtained by means of shaving and abrading the skin with emery paper following cleaning the skin with alcohol. In order to record motion artifacts caused by the oscillation of the platform, electrodes were placed over the right patella in line with the lower limb (Miles et al., 1982). Three layers of adhesive tape between the skin and the bipolar electrodes (10-mm diameter, Contrôle Graphique Medical, Brie-Comte-Robert, France) served as isolation of the muscle activity from the electrodes. In order to assure exclusive artifact recordings, participants were asked to perform a maximal voluntary knee extension against the fixed lever arm of a leg-extension machine. During the contraction, the patella signal remained at baseline levels. Hence, we concluded that non-baseline patella signal recordings during WBV would be due to the shaking of the electrodes and not due to crosstalk from surrounding muscles. The patella and the sEMG signals were amplified (MP150 Biopac® Systems Inc., Holliston, MA, USA; CMRR = 110 db, Z input = 1000 M™¦, gain = 1000), filtered with a bandwidth frequency ranging from 10 Hz to 500 Hz, and online digitized with a sampling frequency of 2000 Hz.
Signal processingIn order to calculate the RMS of the first spike (fundamental) and the following spikes (harmonics), the spikes were separated from the sEMG signal in the frequency domain. This separation of the spikes was performed by using spectral linear interpolation. More specifically, the sEMG signals were first transformed into the Power Spectral Density (PSD). This was accomplished with the help of the Welch method, using Hamming windows with the length of L = 1024. The exact vibration frequency was verified with the help of the acceleration signal from the platform, as peak vibration frequency was identical for the acceleration and the sEMG signals. After locating the spikes in the sEMG signals, their peaks were replaced by a straight line. This filtering regime allowed calculation of the RMS of the interpolated sEMG, the isolated fundamental, as well as the isolated harmonics (Figure 1). The RMS was calculated via the formula:
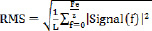
where │Signal(f)│2 was the PSD of the signal, L was the length of the data to compute the PSD, and Fe was the sampling frequency. In order to enable comparison between sEMG signals of the muscles and the electrical signal of the patella, the RMS of the fundamental and the RMS of the harmonics was computed likewise for the electrical signal of the patella. For the patella as well as for the sEMG signals, the ratio between the RMS of the fundamental and the RMS of the harmonics was calculated via the formula:

If γ > 0, the RMS of the fundamental would be higher than the RMS of the harmonics. If γ > γ < 0, the RMS of the harmonics would be higher than the RMS of the fundamental. The γ values of the VL, RF, and BF were averaged to provide a general ratio for the thigh muscles, and the γ values of the SOL and GL were averaged to report the ratio for the calf muscles.
Statistical analysisAll analyses were executed using Statistica software (Statsoft, version 7.0 Tulsa, OK, USA). The Kolmogorov-Smirnov test confirmed the normality of all data (D < 0.24, p > 0.20), and the level of significance was set to α = 0.05. In order to evaluate significant differences between the RMS of the two loads, dependent t-tests were computed. This statistical test was performed for the sEMG of the VL, RF, BF, SOL, and GL, and for the electrical signal of the patella with and without WBV. In order to test if the magnitude of the fundamental and the harmonics increases with increasing sEMG without the spikes, the RMS of the fundamental and the harmonics were correlated to the interpolated RMS of each muscle, calculating Pearson’s correlation coeffficients. For the patella signal, Pearson’s correlation coeffficients were computed for the RMS of the fundamental and the harmonics with the interpolated RMS of all muscles. Data of the two load conditions were combined for the correlation analysis. Unless specified, the results are presented using 95% confidence intervals (CI).
Patella signalThe electrical signal recorded on the patella during WBV consisted of a sinusoidal wave, similarly to the acceleration signal measured on the platform (Figure 2). In the frequency spectrum, excessive peaks were observed in the patella signal at the vibration frequency and to a lesser extent at the multiple harmonics. No significant difference was found between the RMS of the two loads (p > 0.05, Figure 3A) with and without WBV. During WBV, the ratio between the RMS of the fundamental and the RMS of the harmonics was 0.7, averaged for both load conditions.
sEMG signalDuring WBV, the RMS of the fundamental as well as the RMS of the harmonics was significantly higher with the additional load as compared without the load in all the measured muscles (p < 0.05, Table 1). No significant difference between the two loads was found during the no-vibration trial (p > 0.05). Figure 3B illustrates the increase in RMS with the additional load for the VL. The ratios between the RMS of the fundamental and the RMS of the harmonics were = 0.3 for the thigh muscles and = -0.2 for the calf muscles (averaged for both loads).
CorrelationsFor all muscles, the RMS of the fundamental and the RMS of the harmonics demonstrated significant correlations with the interpolated RMS of the respective muscle (p < 0.05). The correlation coeffficient (r) between the fundamental and the interpolated sEMG was 0.76 for the VL (p < 0.001), 0.72 for the RF (p < 0.001), 0.79 for the BF (p < 0.001), 0.89 for the SOL (p < 0.001), and 0.56 for the GL (p = 0.016). For the harmonics, the correlation coeffficient (r) was 0.85 for the VL (p < 0.001), 0.54 for the RF (p = 0.021), 0.81 for the BF (p < 0.001), 0.89 for the SOL (p < 0.001), and 0.92 for the GL (p < 0.001). The RMS of the patella was not related to the RMS of the interpolated sEMG of any muscle that was measured (p > 0.05).
This study aimed to clarify the content of the isolated spikes that are observed in the sEMG spectrum during WBV. The main findings were that (1) sEMG signals during WBV are contaminated by vibration induced motion artifacts, which was confirmed by motion artifact-recordings form the patella. In addition, (2) recordings of motion artifacts was more prevalent in the first spike (fundamental) than in the following spikes (harmonics), as the averaged ratio  for the patella signal was above zero (γ = 0.7). Further it was shown that (3) sEMG signals during WBV may include muscle activity phase-locked to the vibration frequency and its multiple harmonics, because the RMS of the spikes increased with increasing background activation of the muscle and was highly correlated to the RMS of the sEMG signal without the spikes. for the patella signal was above zero (γ = 0.7). Further it was shown that (3) sEMG signals during WBV may include muscle activity phase-locked to the vibration frequency and its multiple harmonics, because the RMS of the spikes increased with increasing background activation of the muscle and was highly correlated to the RMS of the sEMG signal without the spikes. The high peaks observed in the power spectrum of the patella signal showed that motion artifacts contribute to the sEMG signal during WBV, which confirms the first hypothesis. Furthermore, the RMS of the patella’s fundamental was 3.6 times greater than the RMS of the harmonics, which afffirms the second hypothesis. Therefore it can be concluded that sEMG signals measured during WBV are contaminated by motion-induced artifacts, and that the artifacts are more distinct in the fundamental than in the harmonics. Relevance of motion artifacts during WBV using dummy electrodes placed on the patella (Fratini et al., 2009) or the muscle (Ritzmann et al., 2010; Sebik et al., 2013) has been verified before, reporting conflicting results. Fratini et al. (2009) and Sebik et al. ( 2013) proved the existence of motion artifacts by finding excessive peaks at the vibration frequency and its multiple harmonics, whereas Ritzmann et al. (2010) reported a marginal contribution. Possible reasons for the observed discrapancy could be the type of the used equipment such as the electrodes or the cables (Webster, 1984). Using conjoined electrodes vs. single electrodes might have the advantage that the conjoined electrodes are moving in phase to each other, which impedes a difference in potential between the electrodes. Additionally, the degree of cable shielding could have an influence, as motion artifacts are reduced by using shielded cables vs. unshielded cables (Clancy et al., 2002). Together with the findings of the present study it is challenging to acertain why Ritzmann et al. (2010) did not observe significant spikes in the dummy electrode signal. In order to minimize motion artifacts during WBV exercises, the electrodes and their cables should be taped to the skin, the amplifier/preamplifier should be placed close to the recording electrodes, and short cables should be used connecting the electrodes with the amplifier (Türker, 1993). In case recordings of motion artifacts cannot be prevented, they can be removed offfline by deleting the excessive spikes in the sEMG spectrum using a band-stop filter (Abercromby et al., 2007; Fratini et al., 2009; Pollock et al., 2010) or spectral linear interpolation centred at the vibration frequency and its multiple harmonics, or by deleting the entire frequency spectrum where the artifacts might have occurred (Hazell et al., 2007; 2010; Sebik et al., 2013). Deleting the excessive spikes in the sEMG spectrum might not only remove the artifacts, but also parts of the reflex activity evoked by the vibration. This reasoning could explain why numerous authors neglected to filter the sEMG signals during WBV exercises (Di Giminiani et al., 2013; Krol et al., 2011; Perchthaler et al., 2013). It needs to be considered that the motion artifacts are exclusively present in the sharp spikes of the sEMG spectrum (Abercromby et al., 2007; Fratini et al., 2009; Sebik et al., 2013), whereas the reflex activity is spread over a wide frequency range (Ritzmann et al., 2010). Therefore, withdrawal of the spikes in the sEMG spectrum does not affect most of the information that is related to the reflex activity. This characteristic would favour the removal of the excessive spikes in the sEMG spectrum. Interestingly, it was found that the RMS of the fundamental and the RMS of the harmonics increased with increasing background muscle activity. Identical behavior has been reported for stretch reflex responses evoked by directly-applied vibration to the tendon or the muscle in numerous studies (Bedingham and Tatton, 1984; Bongiovanni and Hagbarth, 1990; Burke et al., 1978; 1976a; 1976b; Eklund and Hagbarth, 1966; Nordin and Hagbarth, 1996). Further, there is recent evidence that this mechanism also occurs during WBV exercises (Zaidell et al., 2013). Therefore it is possible that the spikes observed in the sEMG spectrum during WBV consist partially of stretch reflex responses. It is important to mention that we did not measure stretch reflex responses per se. Therefore, other reflex mechanisms induced by e.g. golgi tendon organs and cutaneous mechanoreceptors could explain the findings of the present study. However, these results indicate that the isolated spikes in the sEMG spectrum contain motion artifacts and possibly reflex activity. A higher ratio ratio 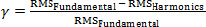 for the patella signal (γ = 0.7) than for the muscles (thigh: γ = 0.3; calf: γ = -0.2) was observed. ratios higher than zero indicate that the RMS of the fundamental was higher than the RMS of the harmonics and vice versa. As the γ ratios were lower in the muscles than in the patella, we conclude that the magnitude of the harmonics compared to the fundamental was higher in the muscles’ sEMG signals than in the patella signal. This observation might be due to contribution of reflex activity in the muscle sEMG. Stretch reflex trains separated by a time interval of t0result in a spectrogram with peaks separated by the frequency interval of for the patella signal (γ = 0.7) than for the muscles (thigh: γ = 0.3; calf: γ = -0.2) was observed. ratios higher than zero indicate that the RMS of the fundamental was higher than the RMS of the harmonics and vice versa. As the γ ratios were lower in the muscles than in the patella, we conclude that the magnitude of the harmonics compared to the fundamental was higher in the muscles’ sEMG signals than in the patella signal. This observation might be due to contribution of reflex activity in the muscle sEMG. Stretch reflex trains separated by a time interval of t0result in a spectrogram with peaks separated by the frequency interval of 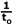 , as illustrated by Ritzmann et al. (2010). However, if such a stretch reflex train is contaminated by motion artifacts, the relative contribution of the fundamental is increased. This effect is shown in Figure 4 with the help of computer simulated signals. Stretch reflex trains were modelled by repeating a simulated signal in the shape of single motor unit action potentials (MUAPS). Although stretch reflex responses of the human body vary in magnitude and time delay (Ritzmann et al., 2010), MUAPS with identical amplitudes and separated by identical time intervals of t0were replicated for simplicity reasons. Figure 4A illustrates the simulated signal of a stretch reflex train in the time domain (upper panel) and the PSD (lower panel) with = 30Hz. In Figure 4B, a sinusoidal wave representing motion artifacts with the same frequency as the stretch reflex train was added to the signal from Figure 4B. Comparing the PSD of the two signals, the ratio was found to be higher in the motion artifact contaminated signal (γ = 0.45) than in the pure stretch reflex train (γ= -0.30). Hence, it can be concluded that the higher the ratio γ, the higher the contribution of motion artifacts and the lower the involvement of reflex activity. The fact that γ was lower for the muscles than the patella further suggests reflex activity in sEMG recordings during WBV exercises. , as illustrated by Ritzmann et al. (2010). However, if such a stretch reflex train is contaminated by motion artifacts, the relative contribution of the fundamental is increased. This effect is shown in Figure 4 with the help of computer simulated signals. Stretch reflex trains were modelled by repeating a simulated signal in the shape of single motor unit action potentials (MUAPS). Although stretch reflex responses of the human body vary in magnitude and time delay (Ritzmann et al., 2010), MUAPS with identical amplitudes and separated by identical time intervals of t0were replicated for simplicity reasons. Figure 4A illustrates the simulated signal of a stretch reflex train in the time domain (upper panel) and the PSD (lower panel) with = 30Hz. In Figure 4B, a sinusoidal wave representing motion artifacts with the same frequency as the stretch reflex train was added to the signal from Figure 4B. Comparing the PSD of the two signals, the ratio was found to be higher in the motion artifact contaminated signal (γ = 0.45) than in the pure stretch reflex train (γ= -0.30). Hence, it can be concluded that the higher the ratio γ, the higher the contribution of motion artifacts and the lower the involvement of reflex activity. The fact that γ was lower for the muscles than the patella further suggests reflex activity in sEMG recordings during WBV exercises. Furthermore, we observed differences in γ ratios between muscle groups, as was lower in γ the calf than the thigh muscles. This indicates that the calf muscles contained higher power in the harmonics compared to the fundamental than the thigh muscles. As the artifacts are showed enhanced reflex activity as compared to the thigh muscles. It is rather unlikely that the sEMG of the calf muscles was less contaminated by motion artifacts as the calf muscles are closer to the vibration platform than the thigh muscles. As previously shown (Fratini et al., 2009), the magnitude of the motion artifacts is a direct result of the magnitude of the local acceleration. Typically, the muscles closer to the platform experience a higher acceleration compared to the muscles further away (Cook et al., 2011). Therefore, it is expected that the sEMG of the calf muscles would be more contaminated by artifact-related recordings. However, the lower γ ratios in the calf muscles could be explained by increased reflex activity (Cochrane et al., 2009) and/or increased muscle damping (Wakeling et al., 2002) as the calf muscles experience higher accelerations due to their proximity to the platform. This could further explain why it has been previously found that the muscles closer to the platform were more affected by WBV than the muscles further away (Pollock et al., 2010; Roelants et al., 2006). One limitation of the present study is the methodological approach of measuring the motion artifacts. The insulating tape placed between the electrodes and the skin indeed prevented possible sEMG recordings from sourrounding muscles, but at the same time elevated the resistance between the electrodes to almost infinite. In order to address this issue, the electrodes could have been connected to a resistor matching the skin impedance to prevent an open circuit (Ritzmann et al., 2010; Sebik et al., 2013). As the use of such a resistor was neglected in the current study set-up, it needs to be acknowledged that the magnitude of the motion artifacts might have been overestimated. Nevertheless, the correlation values (r) as well as the γ ratios are expected to be unchanged despite the absence of the resistor. However, this set-up presents some draw backs even with the use of a resistor. While the artifacts occurring from the cables are included (Clancy et al., 2002; Türker 1993; Webster 1984), possible interference from electrochemical potentials at the electrode/gel interface, at the gel/skin interface, and between skin layers are neglected (Bifulco et al., 2013). Thus, measuring motion artifacts during WBV exercises is faced by limitations which need to be addressed in future studies. The isolated spikes that can be observed in the sEMG spectrum during WBV exercises contain motion artifacts and possibly reflex activity. This implies that the muscle activity measured during WBV exercises can be overestimated when the excessive spikes are not deleted and vice versa. However, further investigations focusing on this topic are needed with separate analysis for the fundamental and the harmonics as the outcomes of this study showed that their contribution in motion artifacts is unequal. |
| AUTHOR BIOGRAPHY |
|
 |
Karin Lienhard |
| Employment: PhD student at the University of Nice Sophia Antipolis, France. |
| Degree: MSc |
| Research interests: Influence of WBV exercises on muscle activity and acceleration transmission |
| E-mail: karin.lienhard@unice.fr |
| |
 |
Aline Cabasson |
| Employment: Associate Professor at the University of Nice Sophia Antipolis, France. |
| Degree: PhD |
| Research interests: Biomedical signal processing, especially in the analysis of the electrocardiograms and electromyograms. |
| E-mail: cabasson@i3s.unice.fr |
| |
 |
Olivier Meste |
| Employment: Professor at the University of Nice Sophia Antipolis, France. |
| Degree: PhD |
| Research interests: Digital processing, time-frequency representations, and modeling of biological signals and systems, including ECG, EMG, and EEG |
| E-mail: olivier.meste@unice.fr |
| |
 |
Serge S. Colson |
| Employment: Professor at the University of Nice Sophia Antipolis, France. |
| Degree: PhD |
| Research interests: Neuromuscular function, especially during fatigue, whole body vibration exercise and strength training programs. |
| E-mail: colson@unice.fr |
| |
|
| |
| REFERENCES |
 Abercromby A.F.J., Amonette W.E., Layne C.S., McFarlin B.K., Hinman M.R., Paloski W.H. (2007) Variation in neuromuscular responses during acute whole-body vibration exercise. Medicine and Science in Sports Exercise 39, 1642-1650. |
Bedingham W., Tatton W.G. (1984) Dependence of EMG responses evoked by imposed wrist displacements on pre-existing activity in the stretched muscles. Canadian Journal of Neurological Sciences 11, 272-280. |
Bifulco P., Cesarelli M., Romano M., Fratini A. (2013) Comments on the article “rectification of SEMG as a tool to demonstrate synchronous motor unit activity during vibration.”. Journal of Electromyography and Kinesiology 23, 1250-1251. |
Bongiovanni L.G., Hagbarth K.E. (1990) Tonic vibration reflexes elicited during fatigue from maximal voluntary contractions in man. Journal of Physiology 423, 1-14. |
Brown M.C., Engberg I., Matthews P.B. (1967) The relative sensitivity to vibration of muscle receptors of the cat. Journal of Physiology 192, 773-800. |
Burke D., Hagbarth K.E., Löfstedt L. (1978) Muscle spindle responses in man to changes in load during accurate position maintenance. Journal of Physiology 276, 159-164. |
Burke D., Hagbarth K.E., Löfstedt L., Wallin B.G. (1976a) The responses of human muscle spindle endings to vibration of non-contracting muscles. Journal of Physiology 261, 673-693. |
Burke D., Hagbarth K.E., Löfstedt L., Wallin B.G. (1976b) The responses of human muscle spindle endings to vibration during isometric contraction. Journal of Physiology 261, 695-711. |
Cardinale M., Bosco C. (2003) The use of vibration as an exercise intervention. Exercise and Sport Sciences Reviews 31, 3-7. |
Cardinale M., Lim J. (2003) Electromyography activity of vastus lateralis muscle during whole-body vibrations of different frequencies. Journal of Strength and Conditioning Research 17, 621-624. |
Clancy E.A., Morin E.L., Merletti R. (2002) Sampling, noise-reduction and amplitude estimation issues in surface electromyography. Journal of Electromyography and Kinesiology 12, 1-16. |
Cochrane D.J., Loram I.D., Stannard S.R., Rittweger J. (2009) Changes in joint angle, muscle-tendon complex length, muscle contractile tissue displacement, and modulation of EMG activity during acute whole-body vibration. Muscle & Nerve 40, 420-429. |
Cook D.P., Mileva K.N., James D.C., Zaidell L.N., Goss V.G., Bowtell J.L. (2011) Triaxial modulation of the acceleration induced in the lower extremity during whole-body vibration training: a pilot study. Journal of Strength and Conditioning Research 25, 298-308. |
Di Giminiani R., Masedu F., Tihanyi J., Scrimaglio R., Valenti M. (2013) The interaction between body position and vibration frequency on acute response to whole body vibration. Journal of Electromyography and Kinesiology 23, 245-251. |
Eklund G., Hagbarth K.E. (1966) Normal variability of tonic vibration reflexes in man. Experimental Neurology 16, 80-92. |
Fratini A., Cesarelli M., Bifulco P., Romano M. (2009) Relevance of motion artifact in electromyography recordings during vibration treatment. Journal of Electromyography and Kinesiology 19, 710-718. |
Hazell T.J., Jakobi J.M., Kenno K.A. (2007) The effects of whole-body vibration on upper- and lower-body EMG during static and dynamic contractions. Applied Physiology, Nutrition and Metabolism 32, 1156-1163. |
Hazell T.J., Kenno K.A., Jakobi J.M. (2010) Evaluation of muscle activity for loaded and unloaded dynamic squats during vertical whole-body vibration. Journal of Strength and Conditioning Research 24, 1860-1865. |
Krol P., Piecha M., Slomka K., Sobota G., Polak A., Juras G. (2011) The effect of whole-body vibration frequency and amplitude on the myoelectric activity of vastus medialis and vastus lateralis. Journal of Sports Science and Medicine 10, 169-174. |
Lienhard K., Cabasson A., Meste O., Colson S.S. (2014) Determination of the optimal parameters maximizing muscle activity of the lower limbs during vertical synchronous whole-body vibration. European Journal of Applied Physiology 114, 1493-1501. |
Lorenzen C., Maschette W., Koh M., Wilson C. (2009) Inconsistent use of terminology in whole body vibration exercise research. Journal of Science and Medicine in Sport 12, 676-678. |
Marín P.J., Santos-Lozano A., Santin-Medeiros F., Vicente-Rodriguez G., Casajús J.A., Hazell T.J., Garatachea N. (2012) Whole-body vibration increases upper and lower body muscle activity in older adults: potential use of vibration accessories. Journal of Electromyography and Kinesiology 22, 456-462. |
Miles T.S., Browne E., Wilkinson T.M. (1982) Identification of movement artefacts in electromyograph recordings. Electromyography and Clinical Neurophysiology 22, 245-249. |
Nordin M., Hagbarth K.E. (1996) Effects of preceding movements and contractions on the tonic vibration reflex of human finger extensor muscles. Acta Physiologica 156, 435-440. |
Perchthaler D., Horstmann T., Grau S. (2013) Variations in neuromuscular activity of thigh muscles during whole-body vibration in consideration of different biomechanical variables. Journal of Sports Science in Medicine 12, 439-446. |
Pollock R.D., Woledge R.C., Martin F.C., Newham D.J. (2012) Effects of whole body vibration on motor unit recruitment and threshold. Journal of Applied Physiology 112, 388-395. |
Pollock R.D., Woledge R.C., Mills K.R., Martin F.C., Newham D.J. (2010) Muscle activity and acceleration during whole body vibration: effect of frequency and amplitude. Clinical Biomechanics 25, 840-846. |
Ritzmann R., Gollhofer A., Kramer A. (2013) The influence of vibration type, frequency, body position and additional load on the neuromuscular activity during whole body vibration. European Journal of Applied Physiology 113, 1-11. |
Ritzmann R., Kramer A., Gruber M., Gollhofer A., Taube W. (2010) EMG activity during whole body vibration: motion artifacts or stretch reflexes?. European Journal of Applied Physiology 110, 143-151. |
Roelants M., Verschueren S.M.P., Delecluse C., Levin O., Stijnen V. (2006) Whole-body-vibration-induced increase in leg muscle activity during different squat exercises. Journal of Strength and Conditioning Research 20, 124-129. |
Roll J.P., Vedel J.P. (1982) Kinaesthetic role of muscle afferents in man, studied by tendon vibration and microneurography. Experimental Brain Research 47, 177-190. |
Sebik O., Karacan I., Cidem M., Türker K.S. (2013) Rectification of SEMG as a tool to demonstrate synchronous motor unit activity during vibration. Journal of Electromyography and Kinesiology 23, 275-284. |
Torvinen S., Kannu P., Sievänen H., Järvinen T.A.H., Pasanen M., Kontulainen S., Järvinen T.L.N., Järvinen M., Oja P., Vuori I. (2002) Effect of a vibration exposure on muscular performance and body balance. Randomized cross-over study. Clinical Physiology and Functional Imaging 22, 145-152. |
Türker K.S. (1993) Electromyography: some methodological problems and issues. Physical Therapy 73, 698-710. |
Wakeling J.M., Kaya M., Temple G.K., Johnston I.A., Herzog W. (2002) Determining patterns of motor recruitment during locomotion. Journal of Experimental Biology 205, 359-369. |
Wilson A. M., McGuigan M., Webster J.G. (1984) Reducing motion artifacts and interference in biopotential recording. IEEE Transactions on Biomedical Engineering 31, 823-826. |
Zaidell L.N., Mileva K.N., Sumners D.P., Bowtell J.L. (2013) Experimental evidence of the tonic vibration reflex during whole-body vibration of the loaded and unloaded leg. PLoS ONE 8, e85247-. |
|
| |
|
|
|
|






